Entwicklungsgenetik
Kontrolle der räumlichen Laserstrahlformung in der Entwicklung der Multiview-Lichtblattmikroskopie
Die Arbeitsgruppe für Entwicklungsgenetik (Developmental Genetics, DG) untersucht die Regulation des dynamischen Zellverhaltens in der Entwicklung des Drosophila melanogaster-Embryos [1]. Zu den Werkzeugen, die bei diesen Studien eingesetzt werden, gehören molekulare Zellbiologie, Genetik und hochauflösende Bildgebung. Der Drosophila-Embryo hat eine recht große Größe (500 µm Länge, 200 µm Durchmesser), was eine Herausforderung für konventionelle mikroskopische Techniken darstellt. Das übergeordnete Ziel unseres Projekts ist es, ein Mikroskop mit hoher räumlicher und zeitlicher Auflösung zu entwickeln und zu bauen, das auf einer selektiven Mikroskopie-Plattform mit ebener Beleuchtung (SPIM oder Lichtblattmikroskopie) basiert. SPIM basiert auf dem optischen Schneiden durch Beleuchten der Probe mit einem dünnen Lichtblatt orthogonal zur Detektionsachse (Abb. 1a). Die Schnittfähigkeit bei SPIM wird durch die Dicke des Lichtblattes definiert. Ein Schwerpunkt der SPIM-Forschung ist es, ein optimales Lichtblatt zu erzeugen, das ein großes Sichtfeld mit einer hohen räumlichen Auflösung abdeckt. Verschiedene Methoden, einschließlich spezielle räumliche Strahlstrukturen (z. B. Bessel-, Airy-, Gauß-Strahlen) oder Tiling-Verfahren, wurden empfohlen, um dieses Problem zu lösen, aber diese sind für Biologen mit wenig Ausbildung in Optik oder Physik oft schwer anzuwenden. Um optimierte Lichtbögen für spezifische biologische Anwendungen zu erzeugen, haben wir eine Software namens Structured SPIM (SSPIM) entwickelt, die einen Open-Source, benutzerfreundlichen und kompakten Werkzeugkasten für die Strahlformung bietet, der digitale Muster für eine Vielzahl von Strukturen zur Formung optischer Strahlen erzeugen kann [2]. SSPIM kann Gauß-, Bessel- und Airy-Strahlen (Abb. 1b-g) durch einfache Ansteuerung eines Spatial Light Modulators (SLM) erzeugen. SSPIM ist auch in der Lage, Muster für inkohärente und kohärente Array-Strahlbildung und Strahlkacheln anzuweisen (Abb. 2) [2]. SSPIM stellt eine Schlüsselkomponente unserer aktuellen Entwicklung eines Dual-View-Tiling-OpenSPIM-Systems dar, mit dem wir nun in der Lage sind, das gesamte funktionale 3D-Bild des dynamischen Zellverhaltens in der Drosophila-Gastrulation mit subzellulärer Auflösung aufzunehmen.
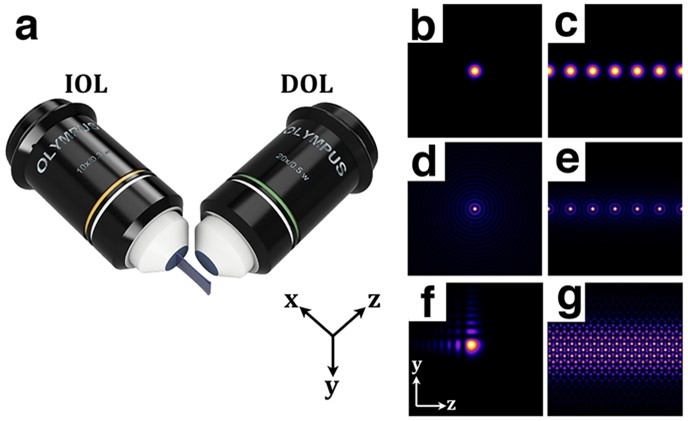
Abbildung 1: Schematische Darstellung des SPIM und Beispiel für Beleuchtungsstrahlen. (a) Prinzip des SPIM: Die Beleuchtungsobjektivlinse (IOL) regt ein dünnes Blatt (x-y-Ebene); das emittierte Licht wird mit dem Detektionsobjektiv (DOL) in der orthogonalen Richtung (z-Achse) gesammelt. (b, d) Transversale Intensitätsprofile von einzelnen Gauß- und Bessel-Strahlen mit radialer Symmetrie und (f) 2D-Aryy-Strahlen mit asymmetrischem Intensitätsprofil. (c,e,g) Für eine bessere Auflösung und einen besseren Kontrast können Gauß- (c) und Bessel-Array-Strahlen (e) und Gitterstrahlen (g) in das SPIM eingesetzt werden
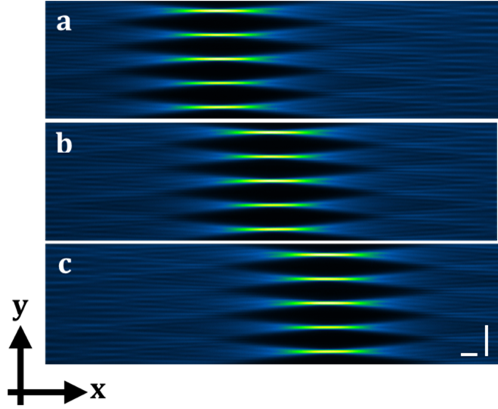
Abbildung 2: Tiling-Methode. (a-c) zeigen das gemessene Intensitätsprofil des gekachelten Gauß-Array-Strahls durch die Farbstofflösung. Die Intensität der grünen Farbe zeigt die dünnsten Lichtschichten und damit die beste räumliche Auflösung an. Die Tiling-Methode ermöglicht die sequentielle Beleuchtung der Probe mit der besten räumlichen Auflösung der Lichtschichten. Die Kachelung wird durch die Anwendung von SSPIM zur Steuerung eines räumlichen Lichtmodulators (SLM) erreicht. Skalenbalken: 40 μm.
Airyscan superauflösende konfokale Mikroskopie zeigt die Dynamik von Zellteilungsproteinen
Ein zweites Forschungsthema in der DG-Gruppe ist die Analyse von Adaptorproteinen des Motorproteins Dynein und deren Rolle bei der Kontrolle der Spezifität und Aktivität des Dynein-vermittelten Transports entlang von Mikrotubuli. In einer aktuellen Studie haben wir die Rolle des Dynein-Adaptors Spindly untersucht, der ursprünglich als ein spezifischer Regulator der Dynein-Aktivität während der Zellteilung identifiziert wurde. Während einige Details der Spindly-Funktion bei Zellteilungen in anderen Systemen herausgearbeitet wurden, war die Funktion von Spindly im Kontext eines Organismus noch nicht untersucht worden. Durch Knock-down des Spindly-Proteins mittels RNA-Interferenz (RNAi) konnte gezeigt werden, dass Spindly für mitotische Zellteilungen im Embryo und für die Zellmigration von Follikelzellen im Eierstock benötigt wird [3]. Dieser genetische Hintergrund wurde genutzt, um die Lokalisation von Spindly mit Hilfe von Transgenen, mit grün fluoreszierendem Protein (GFP) markierten Proteinen zu untersuchen. Wir zeigten zum ersten Mal die dynamische Verteilung des Spindly-Proteins in einem lebenden Organismus (Abb. 3). Das Spindly-GFP-System ermöglichte die Analyse der Spindly-Proteindomänen für seine Lokalisierung und Funktion und zeigte, dass die carboxyterminale Region (DTR) die Spindly-Lokalisierung auf zelltypspezifische Weise kontrolliert. Während die CTR für die Lokalisation und Funktion von Spindly im Embryo entbehrlich ist, ist sie in larvalen Gründerzellen des Nervensystems für die Lokalisation von Spindly notwendig. Wir entdeckten auch, dass eine Erhöhung des Spindly-Spiegels in der weiblichen Keimbahn zu Letalität führt und die Morphologie der Eizelle verändert. Um festzustellen, ob Spindly eine Rolle in post-mitotischen Zellen spielt, veränderten wir den Spindly-Proteinspiegel in migrierenden Zellen und fanden heraus, dass die Migration von Ovarialrandzellen empfindlich auf den Spiegel des Spindly-Proteins reagiert. Zusammenfassend lässt sich sagen, dass unsere Studie den ersten Nachweis der Funktion und der dynamischen Verteilung von Spindly in einem lebenden Organismus liefert und neue Funktionen des Proteins aufdeckt, die eine differenzierte, funktionelle Anforderung für seine carboxy-terminale Region in Drosophila offenbaren.

Referenzen
[1] Winklbauer R and Muller HA (2011). Mesoderm layer formation in Xenopus and Drosophila gastrulation. Phys Biol 8(4):045001.
[2] Aakhte, M, Akhlaghi, EA and HAJ Müller (2018). SSPIM: a beam shaping toolbox for structured selective plane illumination microscopy. Sci. Reports 8, 10067.
[3] Clemente GD, Hannaford MR, Beati H, KappK, Januschke J, Griffis ER and Müller HAJ (2018). Requirements of the Dynein-adaptor Spindly for Mitotic and Post-Mitotic Functions in Drosophila. J Dev Biol. 6 (2), 9.
Kontakt
Prof. Dr. Arno Müller
Ordentliches Mitglied

- Telefon
- +49 561 804-4725
- h.a.muller[at]uni-kassel[dot]de
- Standort
- Universität Kassel
Fachbereich 10 - Naturwissenschaften & Mathematik
Institut für Biologie
Heinrich-Plett-Straße 40
34132 Kassel
- Raum
- IBC, 2403